
 Introduction to Antibiotic Resistance
Introduction to Antibiotic Resistance
Antimicrobial compounds, including natural and synthetic antibiotics, have been crucial in combating infections. Antibiotic resistance, however, has risen rapidly, threatening public health. Resistance can develop through mutations or acquisition of resistance genes via horizontal gene transfer, which has become the primary driver of the current antimicrobial resistance pandemic.
Origins of Antibiotic Resistance
Antibiotic resistance is ancient, arising from interactions between organisms and their environment. Many antibiotic-producing bacteria, such as Streptomyces species, carry self-resistance mechanisms. Environmental non-antibiotic-producing bacteria have also evolved resistance to survive alongside these producers. Even ancient permafrost samples reveal genes resistant to β-lactams, tetracyclines, and glycopeptides, showing the long-standing presence of resistance genes.
(https://share.google/images/BHfaQxPqmbpyS3GPC)
Mechanisms of Horizontal Gene Transfer
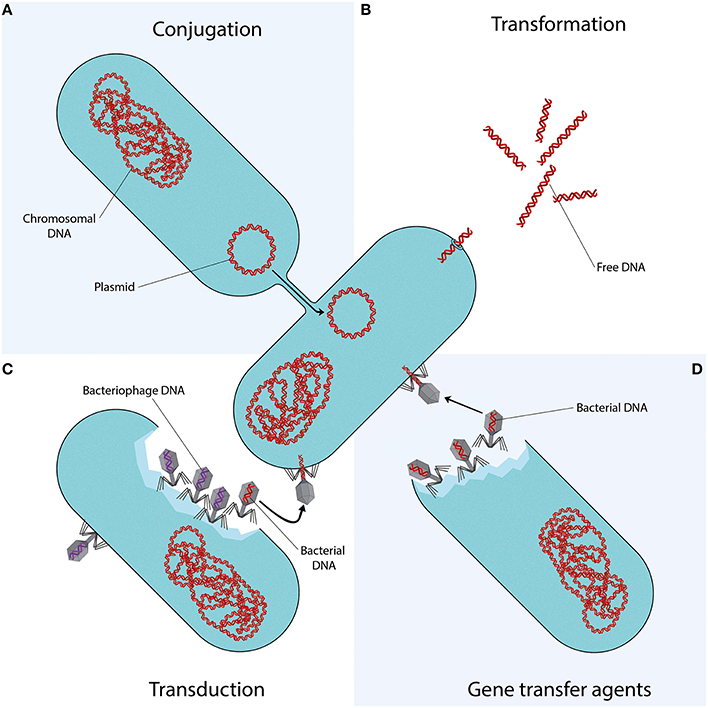
Conjugation
Conjugation involves direct transfer of DNA from one bacterium to another, often via plasmids. Plasmids carrying mobile genetic elements such as transposons or integrons spread resistance genes across bacterial populations, including clinically important genes like blaCTX-M and quinolone resistance genes.
Transformation
In transformation, bacteria uptake DNA fragments from their environment. For instance, penicillin and streptomycin resistance genes were transferred between Streptococcus pneumoniae strains in early experiments, demonstrating this mechanism’s role in spreading resistance.
Transduction
Transduction occurs when bacteriophages transfer DNA between bacteria “by accident.” This process contributes to resistance evolution in species like Staphylococcus aureus and other clinically relevant bacteria.
Environmental Factors and Antibiotic Use
Widespread antibiotic use in medicine, agriculture, and aquaculture accelerates resistance by increasing selective pressure. Most antibiotics are excreted unchanged into the environment, creating hotspots for resistance gene transfer. Increased selection pressure has also accelerated horizontal gene transfer and the abundance of resistome elements.
Conclusion
Horizontal gene transfer—including conjugation, transformation, and transduction—is key to spreading antibiotic resistance genes among bacteria. Understanding these mechanisms is critical to combating the rise of resistant pathogens and protecting public health.
To support responsible antibiotic use, Bioguard offers the miniAST Veterinary Antibiotic Susceptibility Test Analyzer, a tool designed to help combat antimicrobial resistance with game-changing features:
| Feature | Benefit |
| Fast Results | Get results in just 6 hours, enabling swift and confident treatment. |
| Automated Interpretations | Instantly deliver precise susceptibility profiles, supporting faster, more informed clinical decisions and optimizing patient care. |
| Dual-Sample Testing | Double the efficiency with simultaneous analysis of two samples at once. |
| High Accuracy | Achieve an impressive 92% accuracy rate compared to traditional disc diffusion tests. |
📌 Note for Veterinarians:
The miniAST Veterinary Antibiotic Susceptibility Test Analyzer is available exclusively to licensed veterinary clinics and hospitals.
📩 How to Order miniAST
To purchase miniAST or request a quotation, please contact our sales team or email our customer service:
📧 service@bioguardlabs.com
☎️ Please include your hospital name and contact number so our sales representative can follow up with you directly.